
腹腔巨噬细胞对色氨酸的消耗和吲哚胺的产生
LPS可以诱导褪黑素生成增加
睡眠障碍是现代社会的大问题。即使睡眠的微小变化也会影响健康。特别是肺部疾病患者抱怨睡眠问题,如睡眠障碍和失眠。最近的研究表明,睡眠剥夺(SD)和炎症之间存在联系,然而,其潜在的机制仍不清楚。在目前的研究中,我们研究了褪黑素是否对SD大鼠急性肺部炎症有保护作用。使用改进的多平台水浴剥夺雄性ICR小鼠的睡眠3天。用LPS(5毫克/千克)。在第2天给SD小鼠施用褪黑激素(5 mg/kg)和LPS。将小鼠分为对照组、SD组、LPS组、LPS + SD组和LPS + SD +褪黑素组(每组11只)。在褪黑激素和LPS处理24小时后的第3天杀死小鼠。收集肺组织用于组织学检查和蛋白质分析。丙二醛(MDA)水平测定氧化应激的影响。褪黑素恢复了LPS + SD的体重减轻。组织学检查发现LPS + SD大鼠肺泡损伤伴有炎性细胞浸润。褪黑素显著减轻肺泡损伤。在蛋白质印迹分析中,LPS降低了SD小鼠中Bcl-XL和procaspase-3的水平。与LPS + SD相比,用褪黑激素后,Bcl-XL和procaspase-3的水平增加。LPS处理显示了TUNEL阳性细胞的增加,而褪黑激素阻止了LPS + SD动物中细胞死亡的增加。在脂质过氧化试验中,褪黑素显著降低LPS + SD大鼠升高的MDA水平。我们的结果表明褪黑激素通过抗凋亡和抗氧化作用减轻SD期间的急性肺部炎症。
腹腔巨噬细胞对色氨酸的消耗和吲哚胺的产生
摘要
褪黑素已被证明能调节多种免疫功能,一些作者还阐明了白细胞也能产生吲哚胺。事实上,它似乎参与了一些免疫调节轴,包括与干扰素(IFN)产生有关的轴。因此,我们评估了腹腔分离巨噬细胞的色氨酸消耗率、褪黑素和血清素(5-HT)生成率,以及IFN-α和-γ、LPS和肉豆蔻酸佛波酯(PMA)对这些参数的影响。我们的实验结果表明,从正常大鼠腹腔获得的巨噬细胞在与色氨酸共同培养时表现出5-羟色胺-N-乙酰基转移酶活性的增加,这与在培养基中测定的褪黑素生成增加相对应。这一过程由IFN-α和-γ、PMA、LPS和荷瘤(TB)大鼠血清调节,从而可以推测不同的免疫调节循环通过这些细胞产生的褪黑素/血清素的平衡发挥作用。
关键词: 褪黑素生成,血清素,干扰素
引言
近年来,已经进行了许多实验来识别神经内分泌和免疫系统之间双向通信的机制[1]。这些系统之间的联系依赖于几个免疫因子、激素和共同的受体[2–4]。其中,一个可能的候选者,已经提出了大量证据支持[5–8],是神经激素褪黑素(N-乙酰-5-甲氧色胺)。
褪黑素已被证明可以调节多种免疫功能,如自然杀伤细胞(NK)的细胞毒性[9, 10]、抗体产生[11–13]、淋巴细胞增殖、干扰素-γ(IFN-γ)产生[14]以及辅助性T细胞2型功能[5]。一些实验还证明了褪黑素通过内源性阿片片系统对免疫系统的间接影响,该系统能够控制巨噬细胞和淋巴细胞功能[15, 16],或者通过对骨髓的影响[15, 16]。相反,Finocchiaro和同事们[7]表明外周血单核白细胞(PBML)具有将血清素5-羟色胺(5-HT)代谢为褪黑素的能力,开启了神经内分泌系统中双向通路的可能性。事实上,关于IFN-γ,吲哚胺类似乎是一个免疫调节回路的一部分,因为IFN-γ刺激巨噬细胞和淋巴细胞产生血清素和褪黑素,而这些吲哚胺抑制IFN-γ的合成[17, 18]。
因此,为了更好地了解免疫细胞局部产生褪黑素的机制,我们评估了腹膜腔分离巨噬细胞的色氨酸消耗率以及褪黑素和血清素的产生率,以及IFN-α和-γ、脂多糖(LPS)和肉豆蔻酸佛波酯(PMA)对这些参数的影响。
结果
从Wistar大鼠腹腔获得的巨噬细胞,在色氨酸(500 nM)中孵育1小时后,产生5-HT(141.5±8.7 nmol/h 每mg蛋白)和褪黑素(260.5±16.2 nmol/h 每mg蛋白质;表1)。细胞在LPS、PMA和Walker-256 血清TB大鼠血清中孵育后,褪黑素的产生分别增加了31%、48%和52%,而5-HT、LPS和PMA分泌分别减少了26%、35%和35%(表1)。TB大鼠血浆的存在并没有改变巨噬细胞产生5-HT的能力(表1)。IFN-γ或IFN-α的存在使褪黑素的产生分别减少48%和25%,使5-HT的产生量分别提高108%和41%(表1)。
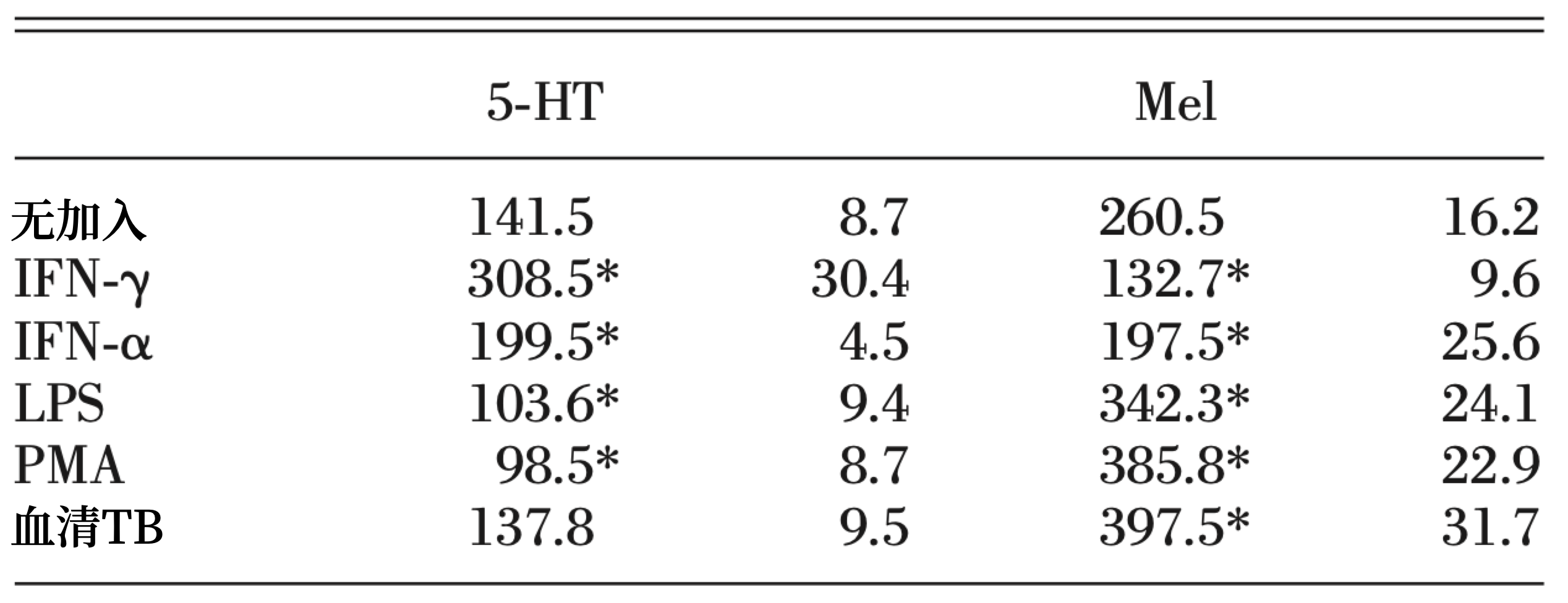
表1 褪黑素和血清素的产生
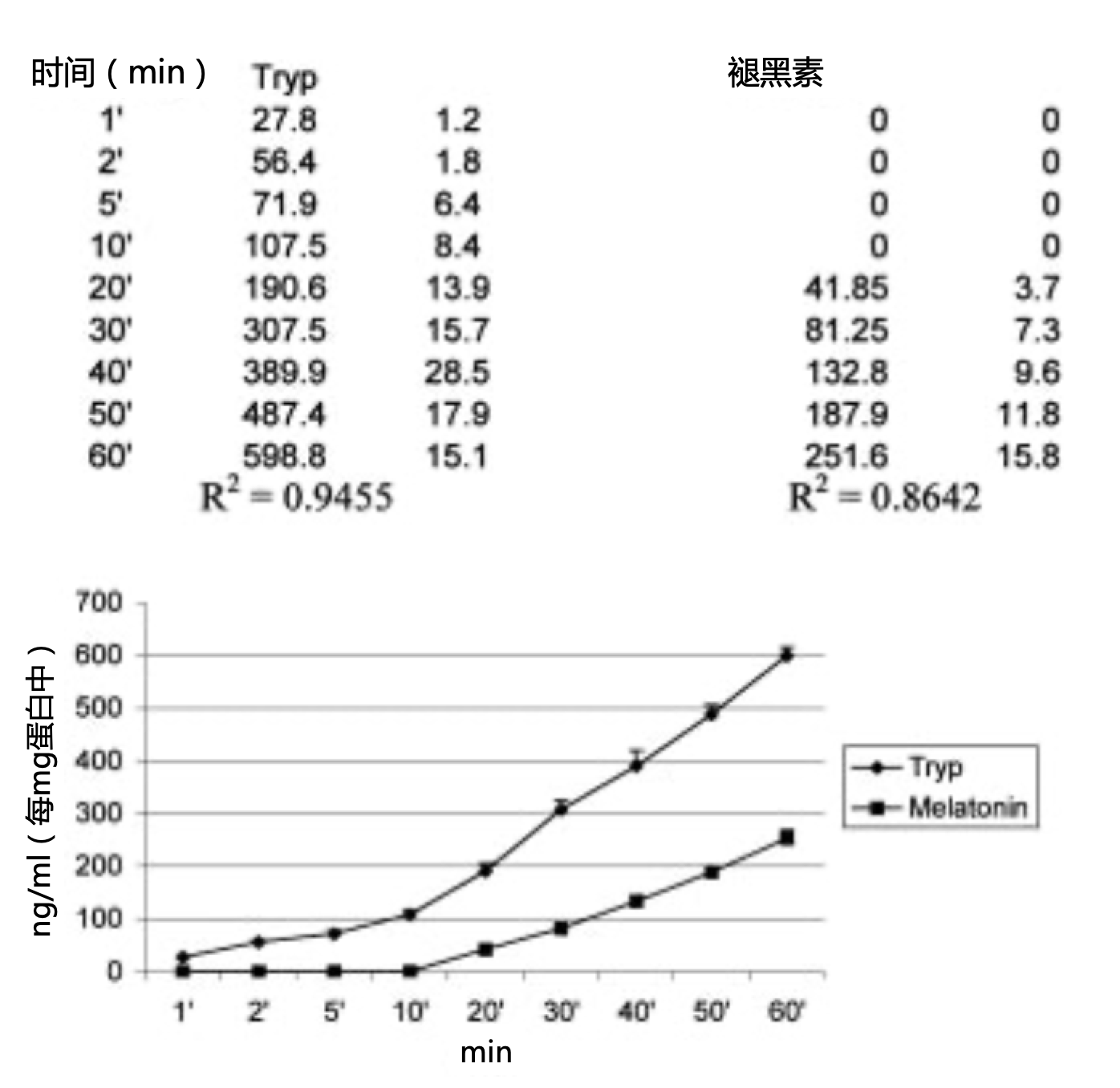
图1. 腹腔巨噬细胞在50 mM色氨酸存在下培养1h后的色氨酸(Tryp)消耗量和褪黑素生成量(ng/mL 每mg蛋白质)。数据表示了五个实验三次重复的平均数±标准差。
变巨噬细胞产生5-HT的能力(表1)。IFN-γ或IFN-α的存在使褪黑素的产生分别减少48%和25%,使5-HT的产生量分别提高108%和41%(表1)。
培养的巨噬细胞在色氨酸消耗开始后20分钟开始产生褪黑素,如图1所示,从该时刻到1小时末呈线性关系(色氨酸消耗量,R2=0.94;褪黑素生成,1小时期间R2=0.86,后40分钟期间R2=0.99;图1)。与图1所示结果相比,向培养基中添加IFN-γ可使色氨酸消耗量增加31%(R2=0.92;图2),并使褪黑素产量减少47%(R2=0.90;图2)。
添加色氨酸(5µM)可将从对照大鼠获得细胞的AA-NAT的最大活性提高11.2倍(表2)。有趣的是,当将PMA添加到培养基中时,未增强对照大鼠细胞中AA-NAT的活性(表2);IFN-γ使酶活性降低了32%(表2),而LPS和TB大鼠血清的酶活性分别提高了28%和21%(表2)。
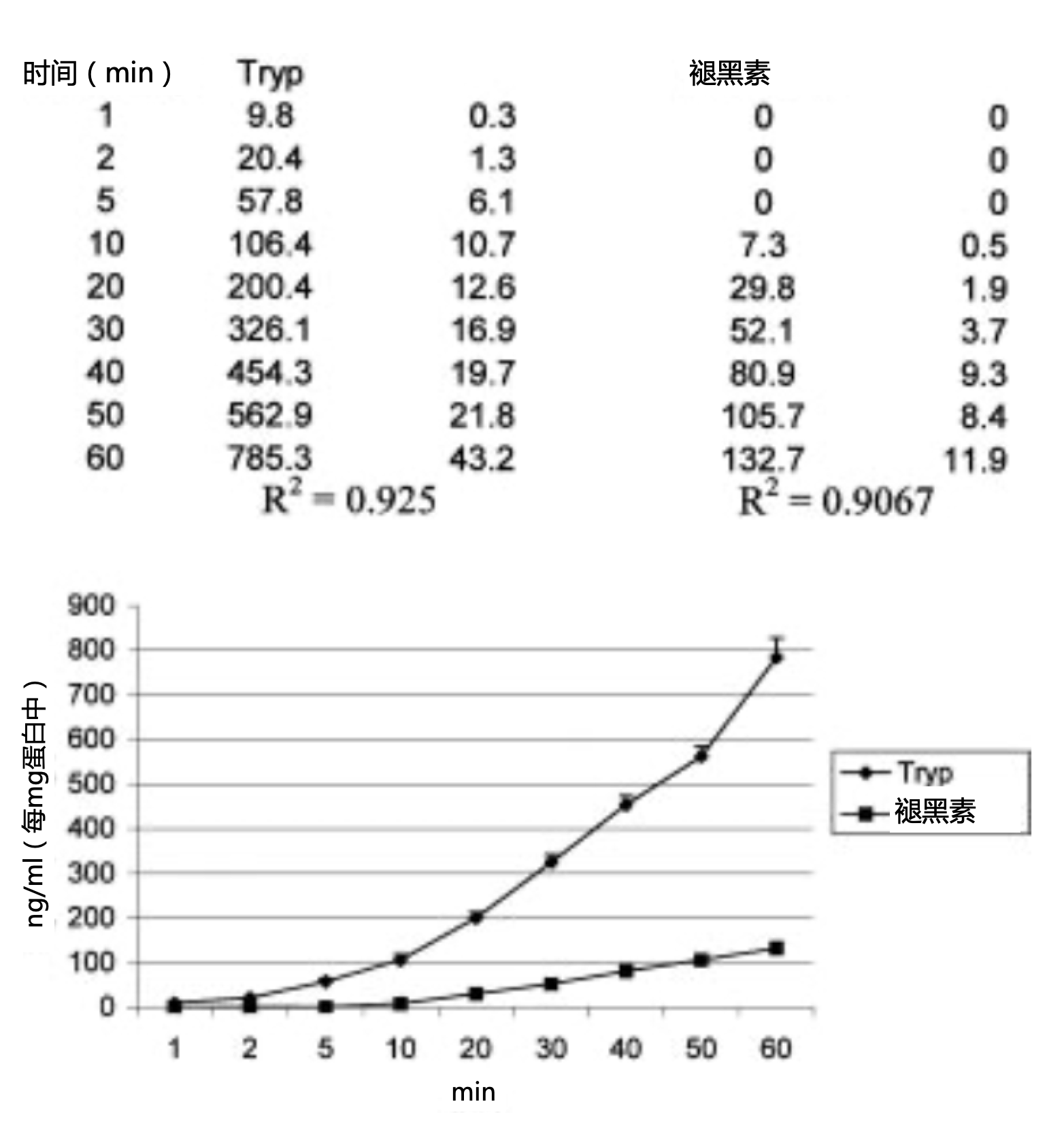
图2. 腹腔巨噬细胞在50 mM色氨酸、10 U/mLIFN-γ存在下培养1h后的色氨酸(Tryp)消耗量和褪黑素生成量(ng/mL 每mg蛋白质)。数据表示了五个实验三次重复的平均值±标准误。
表2. 巨噬细胞中AA-NAT的最大活性

在色氨酸(TryR;50 mM)或色氨酸+PMA(0.8nM)存在下,正常大鼠腹腔巨噬细胞中AA-NAT(NAT)的最大活性。数据以mM/min 每mg蛋白质表示,表示了五个实验的平均值±标准误。*P<0.05与对照组数据比较.05。aP<0.05,用于与Tryp组数值进行比较。
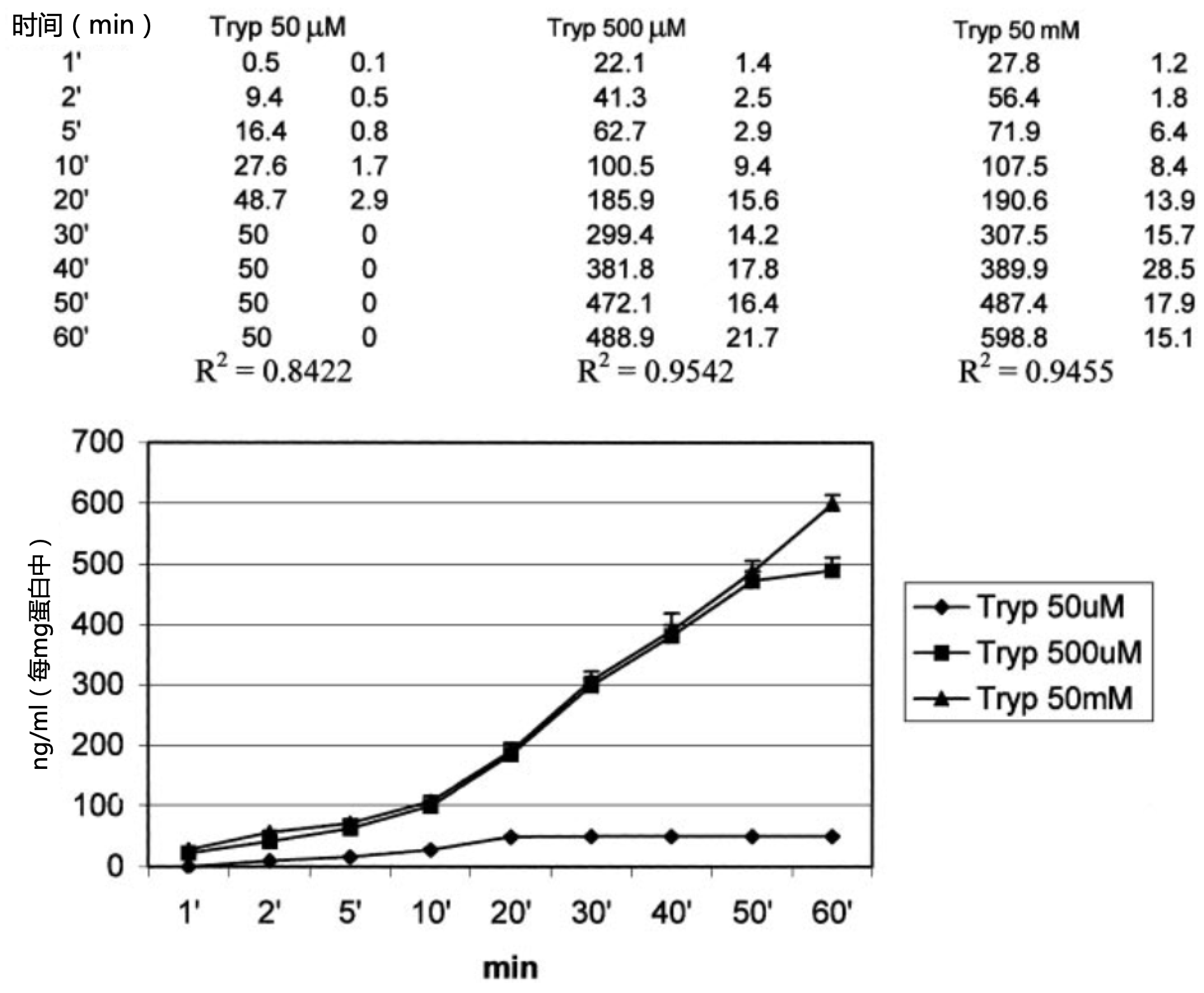
图3. 腹腔巨噬细胞在不同浓度的色氨酸(50 µM、500 µM和5 mM)中培养1h后对色氨酸(Tryp)的消耗。结果以nmol/mg蛋白质表示,数据表示五个重复实验的平均值±标准误。
如图3所示,添加到培养基中的色氨酸量会影响腹腔巨噬细胞的褪黑素产量。事实上,当以生理血浆浓度(50µM,参考文献22)添加色氨酸时,褪黑素的产生在20分钟后达到峰值。500uM色氨酸的存在诱导褪黑素的产生更为显著,在孵育50分钟后达到峰值,在培养60分钟后,50mM色氨酸引起了褪黑素的最大产量(图3)。
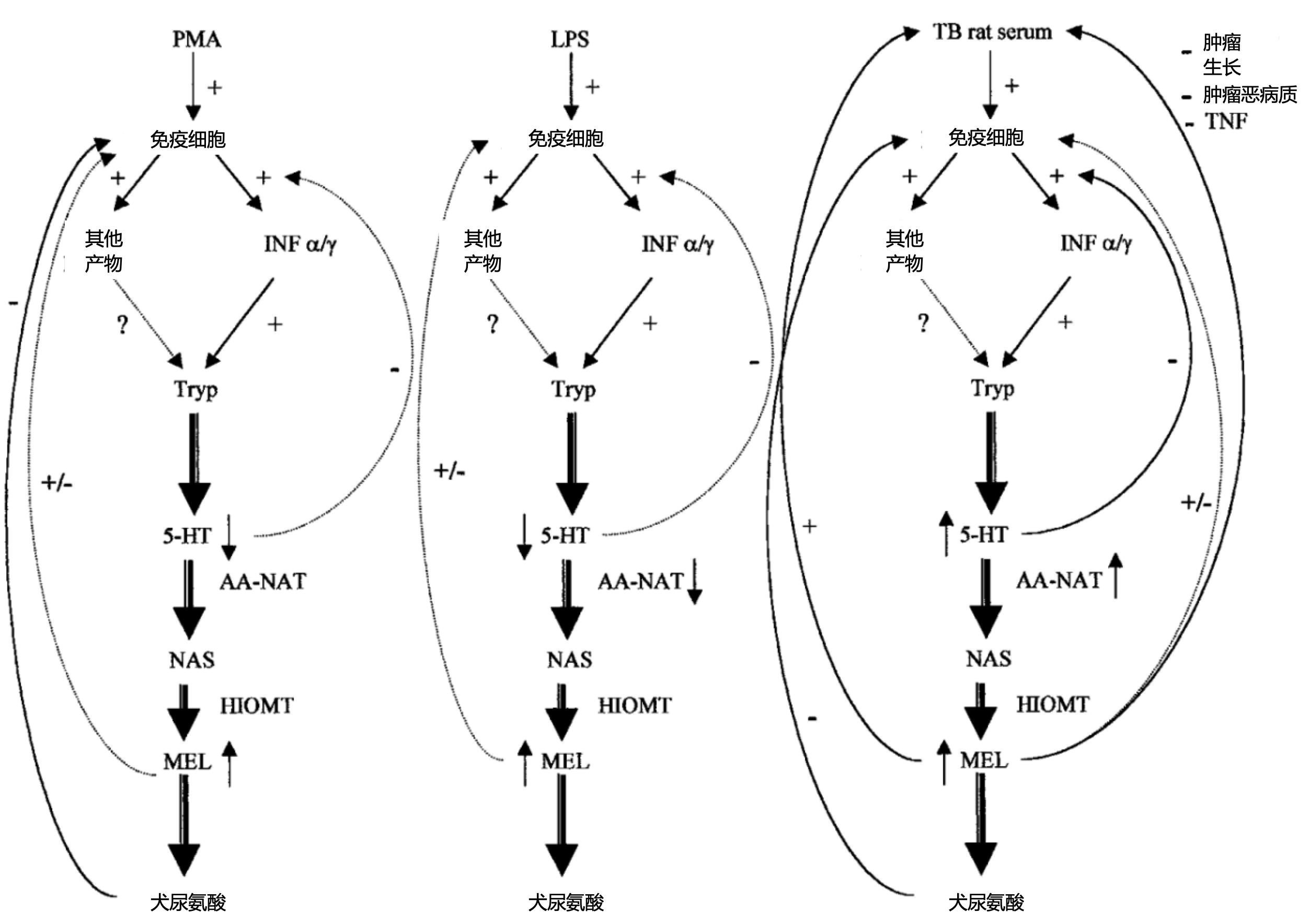
图4. 在LPS、PMA或TB大鼠血清中培养的巨噬细胞可能的调控途径示意图。Tryp:色氨酸;NAS:N-乙酰血清素;MEL:褪黑素;TNF:肿瘤坏死因子;INFɑ/γ:IFN-ɑ/-γ。
讨论
神经内分泌调节因子在免疫系统的发育和功能中起着重要作用,调节适应性和自然免疫[23];免疫细胞产生的许多物质可作为神经内分泌系统[24,25]的反馈信号,其中一些物质也在许多器官和组织中产生,包括中枢神经系统[26]。也很明显,白细胞能够产生经典的激素和神经肽[27]。在参与神经免疫调节的激素中,有大量证据表明,褪黑素作为一个强大的候选人,因为它对神经内分泌系统具有显著影响,是由白细胞产生[5–8],并在维持免疫系统的内稳态方面起着复杂的生理作用[28–30]。PBML产生褪黑素为神经内分泌系统[7]中双向通路的存在提供了可能性。
我们的数据指出,巨噬细胞对色氨酸的消耗量随着氨基酸供给从50 µM增加,在培养30分钟达到峰值时,血浆中氨基酸的生理浓度为50µM20;至50 mM,此时在培养1小时后其我们发现色氨酸消耗呈线性。由于我们在50mM处发现色氨酸呈线性消耗,因此选择这个浓度应用于其他试验。从1小时内提供给细胞的色氨酸总量来看,氨基酸的消耗占培养基中总量的10%,这为其他色氨酸代谢调节机制必要时在其中发挥作用提供可能。
文献中广泛报道了单核细胞源性巨噬细胞消耗色氨酸和释放犬尿氨酸代谢物,作为一种重要的免疫调节机制[31–34],减少色氨酸可用性降低引起T细胞和NK细胞的增殖减少[33,35]以及炎症和免疫介质的产生[31,33,36]。然而,其他色氨酸代谢物,如血清素和相关化合物,也可能以尚未明确[35]的方式影响免疫和炎症反应。
从正常大鼠腹腔获得的巨噬细胞将孵育1小时内消耗的42%的色氨酸转移到褪黑素合成中,从而增强了这些细胞产生吲哚胺的能力,这一点如前Finocchiaro及其同事之前发表的那样[7]。在这些细胞中,褪黑素可以通过产生髓过氧化物酶的犬尿氨酸被氧化,如Rodrigues和同事[37]所述,这为理解色氨酸的免疫调节功能开辟了另一条途径。
考虑到IFN-α和-γ、LPS和PMA刺激白细胞中的色氨酸代谢,导致犬尿氨酸和/或血清素及相关代谢物的生成增加[32,35,38,39],我们评估了这些化合物对与色氨酸孵育的巨噬细胞产生褪黑素和血清素的影响。事实上,我们观察到,这些处理并没有干扰巨噬细胞总体对氨基酸的摄取(约9%),而是显著改变了血清素和褪黑素的生成。IFN-γ减少褪黑素的生成,使色氨酸代谢转向血清素,血清素浓度增加了108%。5-HT浓度的增加可能与AA-NAT和羟基吲哚氧甲基转移酶(HIOMT)的最大活性降低有关,因为这些是褪黑素合成的调节步骤。IFN-α的加入略微增加了血清素的浓度,并降低了褪黑素的浓度,这表明经过AA-NAT和HIOMT的酶的底物流量也减少了。其他的处理,PMA和LPS降低了血清素的浓度,增加了转向褪黑素合成的5-HT的数量。这些结果强烈表明炎症部位存在调节血清素/褪黑素比值的调节机制。
有趣的是,LPS已被证明与褪黑激素协同激活巨噬细胞[40],可以诱导褪黑素生成增加和血清素合成减少。相反,作为Arzt和同事[14]提出的免疫调节回路的一部分,IFN-γ增加了血清素浓度并减少了褪黑素的合成,通过这种回路,IFN-γ刺激的血清素分泌增加以负反馈模式控制这些细胞生成IFN。因此,我们的数据有力地表明,存在与血清素-褪黑素和色氨酸以及细胞因子产生有关的其他免疫调节机制,如图4所示。这些调节机制的存在并不排除通过吲哚胺2,3双加氧酶产生犬尿氨酸的机制,并且与怀孕期间的许多不同情况下的免疫抑制有关[35],在我们的研究中,色氨酸转用于褪黑素和血清素合成的量约占氨基酸总量的10%。
癌症恶病质综合征的临床表现为厌食、消瘦、体重减轻、虚弱、疲劳、表现不佳和免疫功能受损[41]。许多临床和生物学研究表明,该综合征可能与神经内分泌和细胞因子分泌的改变有关。考虑到褪黑素是恶病质期间调节神经免疫内分泌关系的有力候选物质,通过控制神经内分泌和免疫系统[1,5,6]之间的双向通信,我们也评估了TB大鼠血清对腹腔巨噬细胞产生褪黑素和血清素的影响。当巨噬细胞与从TB大鼠获得的血清孵育时,观察到在PMA或LPS存在时发现的相同的反应模式。褪黑素生成的增加可能是另一个免疫调节轴的一部分,因为在晚期癌症患者中[8]褪黑素抑制了TNF的生成,这一反应能减轻恶病质或抑制肿瘤生长[42–44]。相反,肿瘤生长似乎可以调节松果体分泌活性[45]。
考虑到血清素/褪黑素的比值似乎是免疫调节机制的一部分,我们评估了AA-NAT的最大活性,这是褪黑素合成的关键步骤。值得注意的是,当色氨酸添加到培养基中时,AA-NAT的最大活性增加,而PMA的存在,使褪黑激素的生成增加了48%,但并没有改变酶的活性。相反,LPS和TB大鼠的血清增加了酶的最大活性,使更多的色氨酸转向褪黑素的合成中,当IFN-γ添加到培养基中时,酶活性降低了32%。似乎在色氨酸的存在下,酶达到了最大活性,在这种条件下,其他位点如HIOMT可以作为褪黑素合成的控制位点。这些血清素/褪黑素合成途径的可能变化值得更深入地研究。
我们的研究结果表明,从正常大鼠腹腔获得的巨噬细胞,当与色氨酸孵育时,显示AA-NAT活性增加,这与在培养基中测定的褪黑素生成增加相对应。这一过程受IFN-α和-γ、PMA、LPS和TB大鼠血清的调节,从而启发我们探讨不同的免疫调节回路通过这些细胞产生的褪黑素/血清素的平衡发挥作用的可能性。
免责声明
以上部分内容来源于外网我们重在信息分享,版权归属原作者或原刊发平台所有,如有侵权、信息有误或其他异议,请联系我们,我们将在24小时内删除或整改。
复合免疫 健康活力



























