
LPS可触发树突状细胞的成熟和腹腔巨噬细胞的端粒酶活性
LPS可触发树突状细胞的成熟,增强其活性
摘要
端粒酶是维持端粒DNA的逆转录酶,通常在大多数成人组织中检测不到,但在胚胎组织和癌症中呈阳性。此外,新鲜分离或体外激活的淋巴细胞显示出高水平的端粒酶活性,尽管其在包括树突细胞(DCs)在内的骨髓细胞中的表达在很大程度上是未知的。在这里,我们研究了树突状细胞分化和成熟过程中的端粒酶活性。骨髓(BM)细胞与粒细胞-巨噬细胞集落刺激因子和白细胞介素-4的体外培养诱导端粒酶活性显著增加,并伴随其向树突状细胞的分化。此外,用微生物成分如脂多糖(LPS)刺激,可触发树突状细胞的成熟,增强其活性。通过腹腔注射LPS还观察到脾脏树突状细胞中端粒酶活性的体内反应。有趣的是,在老年小鼠脾脏树突状细胞的端粒酶活性明显高于年轻小鼠,但在LPS刺激后反而下降。通过测量细胞表面活化标记物的表达,老年小鼠的脾脏树突状细胞对LPS刺激反应不佳。在骨髓来源的树突状细胞中也观察到了这种对LPS的不良反应。年轻和年老小鼠之间树突状细胞的这些不同特征可能有助于微生物感染的发病机制。
引言
树突状细胞(DC)是一种专业的抗原呈递细胞(APCs ),在原发性免疫反应中起着关键作用[1,2]。树突状细胞在非淋巴组织中处于不成熟状态,在那里它们有效地捕获和处理抗原。激活后,它们启动一个复杂的成熟过程,导致抗原处理能力下降,主要组织相容性复合体(MHC)和共刺激分子的表达增强,并迁移到次级淋巴器官,启动获得性免疫反应[3–6]。成熟过程对树突状细胞的功能至关重要,并使它们能够依次执行不同的、高度专业化的动作。有许多刺激可以在体内和体外启动这一成熟过程。事实上,树突状细胞对许多不同的感染指标都很敏感,这反映了识别多种病原体的关键作用。炎症产物和微生物成分,如脂多糖(LPS)、细菌DNA[CpG-寡脱氧核苷酸(ODN)]和合成的dsRNA (poly I:C),其通常被用作病毒感染的模型,都能够刺激DC变得活化和成熟,专业的APCs[7–10]。
线性真核生物染色体的末端被称为端粒的特殊DNA蛋白质结构所覆盖,端粒由六核苷酸重复序列(TTAGGG)n [11,12]组成。每当体细胞分裂时,端粒DNA就会丢失,这种缩短可能充当有丝分裂时钟,调节细胞分裂的数量。当端粒缩短到一个可能不再稳定染色体末端的临界点时,大多数细胞退出细胞周期并死亡[13,14,15]。端粒酶是一种核糖核蛋白酶,能够将端粒重复序列添加到染色体末端。在端粒酶存在的情况下,端粒长度被延长或维持,并避免了复制性衰老的[16,17]。在最初的分析中,端粒酶活性仅在早期的未成熟细胞中检测到,如婴儿时期骨髓(BM)中的造血前体细胞,但最近的研究表明,在正常的体细胞中可检测到端粒酶活性。已有研究表明,在T细胞激活[18–21]和B细胞分化[22]过程中,端粒酶活性上调,并且在胸腺细胞和生发中心B细胞中观察到高水平的端粒酶活性。然而,有限数量的研究检测了小鼠组织中端粒酶活性的表达,并报道了多种小鼠组织中低水平的活性。
在这里,我们分析了树突状细胞在不同分化阶段的端粒酶活性。在体外和体内诱导的树突状细胞分化成熟过程中,端粒酶活性显著增加。我们还测量了有或没有微生物成分刺激的树突状细胞的端粒酶活性,并在年轻和老年小鼠之间进行了比较。
结果
树突状细胞体外分化过程中端粒酶活性上调
为了探讨树突状细胞分化过程中端粒酶活性是否发生了变化,首先,我们采用了成熟的标准化方法在体外诱导树突状细胞。用GM-CSF和IL-4培养6-8周龄小鼠的BM细胞,生成ImDCs,每2天检查一次端粒酶活性和表面表型(图1A)。细胞的端粒酶活性在培养后立即开始增加,然后出现急剧增加,并在第8天达到最高水平。同时,表明向树突状细胞分化的CD11c阳性细胞的比例也以类似的方式增加(图1A)。
图1
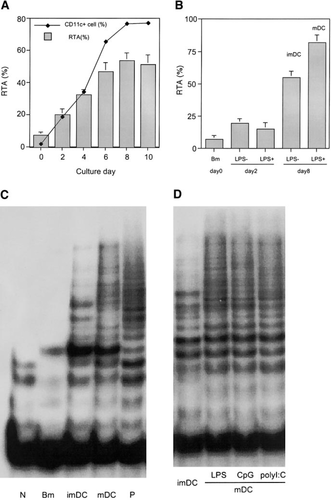
DC分化成熟过程中端粒酶活性的诱导。(A)分离BM细胞并在含有GM-CSF和IL-4的培养基中培养。从第0天到第10天,每2天收集一次细胞,ELISA测量端粒酶活性。相同的细胞用抗CD11c抗体染色并在流式细胞仪中测量。通过同种型对照Ab染色减去CD11c的百分比+细胞是从整个事件计算出来的。(B)在第0天和第6天,收集非粘附细胞,在有或没有LPS的情况下再培养2天,并测量端粒酶活性。(C)通过TRAP观察端粒酶活性。对来自BM(第0天)、imDCs和mDCs(第8天)的PCR产物进行电泳凝胶。(D)在BM培养的第6天,收获细胞并在有或没有CpG-ODN、poly I:C或LPS的情况下再培养2天。用TRAP法比较未受刺激的imDCs和受刺激的mDCs的端粒酶活性。p,阳性对照(提供阳性对照细胞提取物中的端粒酶活性);n,阴性对照(在没有细胞提取物的情况下测量端粒酶活性)。
暴露于LPS等微生物成分可诱导mDC,其特征是共刺激分子的上调。为了评估LPS的作用,我们测量了细胞在有或没有LPS刺激的几个培养时间点的端粒酶活性。虽然我们在第2天没有观察到LPS对端粒酶活性的影响,此时BM细胞还没有很好地分化成DC,但是在第8天用LPS刺激的细胞(mDCs)的端粒酶活性显著高于未刺激的细胞(imDCs),表达中等水平的端粒酶活性(图1B)。在TRAP检测中也观察到相同的结果(图1C)。为了证实端粒酶活性与DC成熟之间的相关性,我们使用了其他微生物来源的炎症因子来激活imDC。最近的研究表明,LPS、CpG和dsRNA能够通过toll样受体(TLR)信号[10,24]触发树突状细胞的成熟。我们的实验表明,与LPS相比,CpG和poly I:C也能提高imdc的端粒酶活性(图1D)。这些结果证明,imdc的端粒酶活性确实在感染因子的成熟过程中被上调。我们在六个独立的实验中都得到了类似的结果。
LPS可增强体内脾脏树突状细胞和腹腔巨噬细胞的端粒酶活性
LPS也被证明可以诱导体内树突状细胞的成熟,而小鼠注射LPS可以诱导树突状细胞的表达上调脾脏树突状细胞[25]的共刺激分子表达。为了进一步探讨端粒酶活性调控与树突状细胞刺激之间的相关性,我们研究了LPS在体内的作用。注射LPS或仅注射PBS的10周龄小鼠的脾细胞通过抗体包被的微珠进行分类:树突状细胞的CD11c珠,单核细胞的单核细胞(巨噬细胞)的CD11b),T细胞的CD90,B细胞的CD19。分选后,用流式细胞术检查细胞,显示其纯度超过90%(数据未显示)。对仅注射PBS的脾细胞的端粒酶活性分析显示,树突状细胞和单核细胞等髓系细胞具有端粒酶活性的中、低水平,淋巴样细胞(B细胞和T细胞)具有较高水平的活性。而注射LPS后,树突状细胞的端粒酶活性显著增强,而其他造血细胞的端粒酶活性仅适度增加(图2A)。综上所述,这些结果表明LPS也能诱导树突状细胞端粒酶活性上调,而树突状细胞是脾脏对LPS刺激的有效反应细胞。在分选的CD11b细胞中,25%细胞也表达代表树突状细胞的CD11c。为了研究其他髓系细胞对LPS的反应,我们还测量了腹腔巨噬细胞的端粒酶活性(图2B)。与树突状细胞相似,腹腔巨噬细胞在无刺激的情况下表达了中等水平的端粒酶活性(50% RTA),并在体内受到LPS刺激后上升到70% RTA。
图2
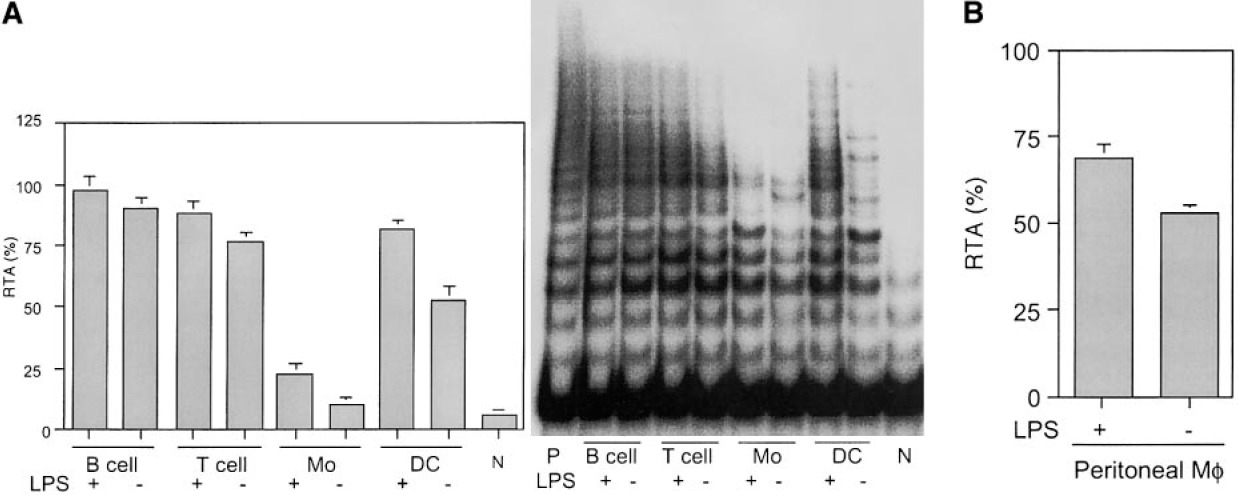
LPS对脾细胞和腹腔巨噬细胞端粒酶活性的影响。(A)将LPS或PBS腹膜内注射入小鼠。48小时后,通过抗体微珠分选脾细胞,并通过抗CD11c将其分离为树突状细胞,通过抗CD11b将其分离为单核细胞(Mo),通过抗CD90将其分离为T细胞,通过抗CD19将其分离为B细胞。ELISA和TRAP检测每个群体的端粒酶活性。p,阳性对照;n,阴性对照。(B)注射LPS或PBS后,ELISA测定腹腔巨噬细胞(M)的端粒酶活性。
幼年和老年小鼠树突状细胞的不同特征
为了研究年龄对端粒酶活性变化的影响,我们比较了青年和老年小鼠脾细胞的端粒酶活性。尽管淋巴样细胞(B细胞和T细胞)的端粒酶活性基本相似,但老年小鼠的端粒酶活性明显高于年轻小鼠(P<0.01;图3A)。此外,来自老年小鼠的脾树突状细胞在体外用LPS刺激后反而降低了端粒酶活性,尽管来自年轻小鼠的树突状细胞显示了约10%的轻微增加。通过FACS分析,通过CD11c、II类和CD86表达测定,来自两代的新鲜分离的脾树突状细胞显示相似的细胞表面表型(图3B).然而,在刺激后,激活标志物如MHC II类和CD86的表达在老年小鼠中明显降低,表明对LPS的反应较差(图3B)。我们还尝试通过注射LPS进行体内刺激,但老年小鼠没有抵抗力并死亡(数据未显示)。
图3
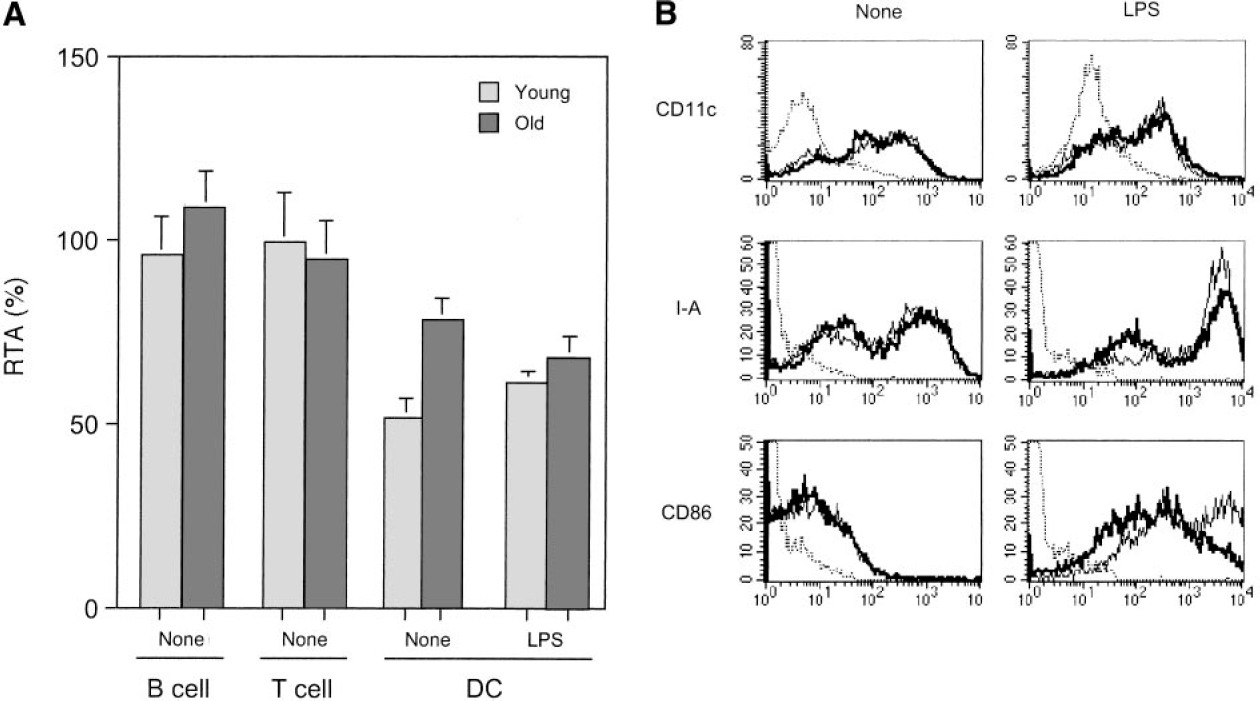
青年和老年小鼠脾脏树突状细胞端粒酶活性和表面表型的比较。(A)在年轻和年老小鼠之间比较新鲜分离的脾B细胞、T细胞、DC和LPS刺激的DC的端粒酶活性。(B)在来自年轻(细线)和年老(粗线)小鼠的新鲜分离的脾树突状细胞和LPS刺激的树突状细胞中,通过荧光素活化的细胞分选仪(FACS)测量CD11c和树突状细胞活化标记[MHC II类(I-A)和CD86]的表达。虚线表示不相关Ab的背景染色。
接下来,我们比较了骨髓树突状细胞的端粒酶活性(图4)。我们在来自幼年小鼠(6周)、成年小鼠(1岁)和老年小鼠(2岁)的BM细胞的同一组体外培养物中产生imDCs。与脾树突状细胞的结果相反,来自老年小鼠的骨髓树突状细胞的端粒酶活性始终低于来自年轻和成年小鼠的树突状细胞,二者具有相似的活性(图4A)。在第6天,用LPS、CpG-ODN或poly I:C刺激每一代的imDCs以诱导成熟。在该实验中,各代小鼠刺激后都观察到了端粒酶活性的上调,但是老年小鼠中的最低活性仍然是一致的(图4B)。通过FACS分析,激活标志物如CD40、CD86和MHC II类的表达在老年小鼠中较低,并且这种趋势在刺激后变得更显著(图4C)。因此,老年小鼠骨髓树突状细胞端粒酶活性降低似乎与激活不足有关。
图4
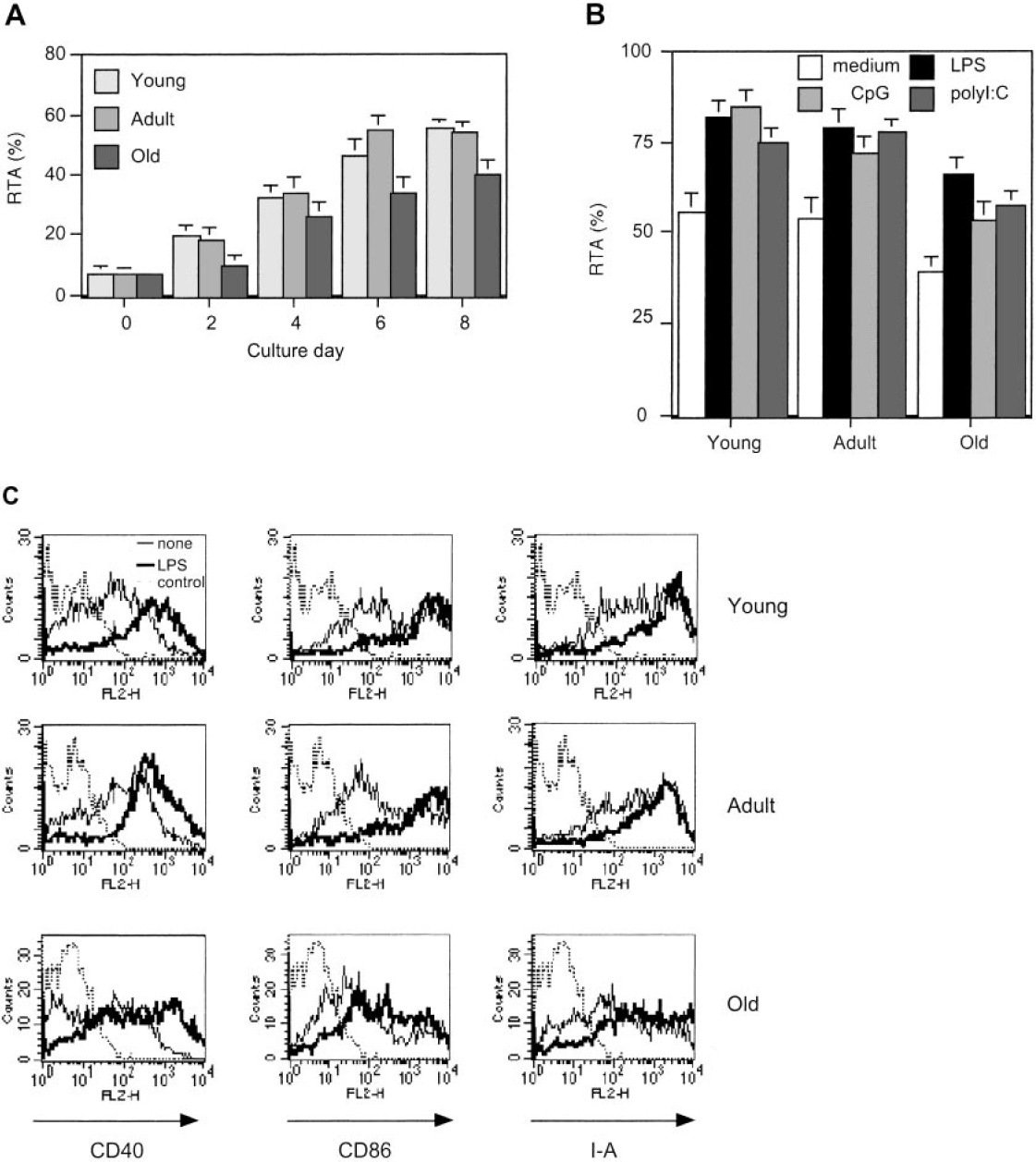
年轻、成年和老年小鼠骨髓树突状细胞的比较。(A)来自老年、成年和年轻小鼠的BM细胞在含有GM-CSF和IL-4的培养基中培养。每隔2天,按照图例中所述分析端粒酶活性图1。(B) ELISA测量每一代用LPS、CpG或poly I:C处理或未处理的DC的端粒酶活性。(C)每一代的imDCs(细线)和LPS诱导的mDCs(粗线)测量DC的活化标记的表达。
讨论
树突状细胞端粒酶
端粒在稳定染色体末端和防止末端融合方面具有重要作用,并且已经发现它的表达与许多不同类型细胞中的细胞增殖相关。因此,端粒酶的激活在决定细胞的增殖能力和抵消端粒的丢失方面具有重要作用。已经表明,T细胞和B细胞活化后,端粒酶活性增加;然而,有限数量的研究显示了髓系细胞的端粒长度和端粒酶活性。据报道,在添加IL-3、flt3配体和干细胞因子的培养物中,成熟髓系祖细胞端粒酶活性上调,在外周血CD34细胞[26,27]肥大细胞发育过程中也诱导端粒酶活性。GM-CSF通过与视黄酸的协同作用抑制髓系白血病细胞中的端粒酶活性和端粒酶逆转录酶(TERT)表达[28]。因此,这引发了一个问题:作为专业的抗原呈递细胞(APC),DCs(树突状细胞)在分化过程中是否会改变端粒酶活性。
在这里,我们清楚地证明了在树突状细胞的分化和成熟过程中,端粒酶活性大幅增加。在GM-CSF和IL-4体外诱导树突状细胞时,BM细胞整个提取物的端粒酶活性开始较低,但在分化成CD11c的过程中逐渐增加+imDCs(图1)。当LPS、CpG-ODN和dsRNA诱导DC成熟时,具有中等水平的imDCs的端粒酶活性在成熟后达到高峰。我们用于DC成熟的所有微生物产品通过TLR途径传导激活信号。TLRs使用相同的信号分子,包括MyD88、IL-1受体相关蛋白激酶和肿瘤坏死因子(TNF)受体相关因子6,导致核因子(NF)-κB的激活[10]。小鼠TERT启动子受广泛表达和高度诱导的NF-κB转录因子调节的事实进一步支持了我们在DC成熟过程中端粒酶激活的发现[29]。然而,诱导型端粒酶活性的生理作用仍不清楚。除了端粒酶在添加端粒重复序列方面的特征性作用之外,应该注意的是,最近的研究表明端粒酶可能具有其他的功能,例如,在保护细胞增殖能力和在没有端粒延伸的情况下防止凋亡方面[30]。实际上,在我们的实验中,在通过微生物成分从imDCs分化为mDCs的过程中,端粒酶活性在没有细胞增殖的情况下增加(数据未显示)。TLRs、CD40L或肿瘤坏死因子相关激活诱导的细胞因子受体的参与通过NF-κB激活延长了树突状细胞的寿命[31,32],其也正向控制端粒酶表达。考虑到最近的研究发现,即LPS和CpG不仅使DC成熟,而且抑制DC凋亡[33,34],端粒酶激活可能有助于这种抗凋亡作用。此外,据报道,树突状细胞是细胞毒性T细胞和NK细胞介导的杀伤的目标,并且由LPS或CD40激活的树突状细胞使树突状细胞对细胞毒性具有抗性[35,36]。综上所述,可诱导的端粒酶活性可能是增强DC功能、增殖和存活的关键因素。
老年小鼠的树突状细胞
尽管最近在树突状细胞的分子和细胞生物学方面取得了许多进展,但很少有研究小组讨论树突状细胞和衰老[37]的话题。在老年小鼠的动物模型中,淋巴结的树突状细胞表现出退行性特征,粘附分子表达减少,树突形成减少,抗原捕获能力降低,这些都意味着其功能活性的破坏[38]。相比之下,从老年人外周血产生的树突状细胞诱导细胞反应的能力没有受损[39]。然而,在衰老过程中,免疫系统的反应性明显下降,这归因于与先天免疫反应相关的淋巴细胞功能受损[40,41]。在这里,我们观察到老年小鼠的脾脏树突状细胞的端粒酶活性远高于年轻小鼠,但在LPS刺激后端粒酶活性降低了(图3A)。相反,来自老年小鼠的BM DCs的端粒酶活性比来自年轻小鼠的低得多,并且在用微生物成分刺激后仍然较弱,同时激活标记物表达较低(图4)。因此,老年小鼠的脾脏树突状细胞和骨髓树突状细胞具有不同的性质。脾脏和骨髓树突状细胞之间这一矛盾数据背后的机制和生理意义仍有待研究。由于老年小鼠中BM DCs的端粒酶活性较低,似乎与激活状态不足有关(图4C),这可能反映了BM细胞的分化和再生潜能较差。相反,老年小鼠的外周脾树突状细胞可能会补偿,如端粒缩短或树突状细胞的功能缺陷,并暗示细胞命运的终末阶段。然而,在脾脏和骨髓树突状细胞中,激活标志物的表达对LPS刺激的不良反应是一致的(图。3B和4C)。在任何情况下,老年小鼠的树突状细胞都具有与年轻小鼠不同的特征,这可能导致其发病机制和对感染或伴随衰老的感染或内毒素休克的抵抗力差。
这是首次报道研究树突状细胞的端粒酶活性,并对年轻和老年小鼠进行比较的报告。我们的观察将为免疫生物学和免疫衰老提供一个新的见解。对端粒酶敲除小鼠树突状细胞的分析(mTERT−/−) [42]将阐明端粒酶在免疫中的生理作用。
免责声明
以上部分内容来源于外网我们重在信息分享,版权归属原作者或原刊发平台所有,如有侵权、信息有误或其他异议,请联系我们,我们将在24小时内删除或整改。
复合免疫 健康活力



























